Research Report
Pathogen Identification of Clubroot Disease in Chinese Cabbage from Yuanyang County, Henan Province 
2 Institute of Horticulture, Henan Academy of Agricultural Sciences, Zhengzhou, Henan, China
* These authors contributed equally to this work
 Author
Author  Correspondence author
Correspondence author
Molecular Plant Breeding, 2017, Vol. 8, No. 5 doi: 10.5376/mpb.2017.08.0005
Received: 28 Feb., 2017 Accepted: 13 Jun., 2017 Published: 28 Jul., 2017
Li Y., Yuan Y.X., Zhao Y.Y., Wei X.C., Yao Q.J., Jiang W.S., Wang Z.Y., Yang S.J., Zhang X.W., and Tian B.M., 2017, Pathogen identification of clubroot disease in Chinese cabbage from Yuanyang county, Henan province, Molecular Plant Breeding, 8(5): 45-51 (doi: 10.5376/mpb.2017.08.0005)
To clarify the causal agent of clubroot disease in Chinese cabbage from Yuanyang experimental base, the isolate YY was collected from the clubroot for further research. After morphological, cytological observation and pathogenicity identification, the pathogen was preliminary inferred as Plasmodiophora brassicae. In order to quickly detect the pathogen, specific primers were designed according to D85819 and Pro1 gene, respectively, and used to amplify the expected fragment from the template DNA of target pathogen. After PCR and sequence analysis, they showed 99% and 93% identity with the D85819 and Pro1 gene sequences of P. brassicae from NCBI respectively. These results indicated that the pathogen was finally confirmed as P. brassicae, which were coincident with preliminary speculation. Moreover, Pro1-specific marker developed in this study can be applied to quick and accurate detection of P. brassicae isolate, which appears a useful assay method for detecting P. brassicae resting spores and improves efforts to halt the dissemination and better control this pathogen.
Background
Plasmodiophora brassicae causes clubroot disease in cruciferous plants by infecting susceptible plant roots, leading to the gall on infected roots as well as leaves wilting, which eventually significantly reduces the quality and production of cruciferous crops (Dixon, 2009; Kim et al., 2009). The clubroot has been one of the most economically devastating disease in Brassica rapa and other cruciferous crops worldwide (Wang et al., 2008). In China, the clubroot disease has spread to the whole country with soil acidification, climate changes, increasing cultivation area of cruciferous crops as well as transportation from south to north and so on (Li et al., 2013; Laila et al., 2017).
P. brassicae is a soil-borne, obligate parasite within the class Phytomyxea (plasmodiophorids) of the protist supergroup Rhizaria (Hwang et al., 2012). Resting spores of P. brassicae released from rot galls have the potential to survive in the soil for up to 20 years (Karling, 1968; Wallenhammar, 1996), which make the disease hard to eradicate. Once clubroot disease occurred, the resting spores in the soil will multiply exponentially, increase disease severity and pose a threat to successive cruciferous crops.
In plant disease diagnostics, many methods have been applied to detect the contamination of P. brassicae in soil. As P. brassicae is an obligate intracellular parasite and hard to culture in vitro, and traditional detection method based on plant bioassays are time-consuming and require large amounts of space, the application of polymerase chain reaction (PCR)-marker based detection has become essential and advantageous for quick and accurate identification of non-culturable obligate parasites (Wallenhammar and Arwidsson, 2001). Many DNA-based methods have all been used to test soil, water, substrates or plant samples for clubroot because of its rapidness, accurateness and convenience (Faggian and Strelkov, 2009). Specific primers for PCR amplification are developed to detect P. brassicae in soil (Ito et al., 1997; 2008; Yang et al., 2002; Yang, 2013). More effective markers should be developed for detection of P. brassicae to prevent any contamination from soil, substrate or plants.
Ribosomal DNA (rDNA) sequence specific marker has usually been usedto detect P. brassicae (Laila et al., 2017; Niwa et al., 2011).A cloned genomic survey sequence of P. brassicae (GenBank accession number D85819; isolate: Hagi) has also been used for the detection of P. brassicae in soil (Ito et al., 1997; Yang, 2013). Pro1 gene from P. brassicae (GenBank accession number GU082362) is a single-copy gene widely present in genomes of P. brassicae, coding serine protease which might play a role during clubroot pathogenesis by stimulating resting spore germination. Therefore it was predicted that Pro1 gene may be an important pathogenicity factor during infection process (Feng et al., 2010).
In March, 2016, a diseased Chinese cabbage plant was found in Yuanyang County, Henan Province, where is an experimental base of Henan Academy of Agricultural Sciences. Its leaves began to wilt, and the root swelled heavily to be a gall. Given the location soil pH value was 8.1 which is obviously beyond the proper range during which clubroot occurs (pH=5.4-6.5) (Li, 2010). In order to figure out the causal agent of infected plant, multiple methods including morphological, cytological observation and pathogenicity identification, combined with molecular biology identification were used in present study. The objective of this study is to accurately confirm and quickly detect the pathogen causing clubroot disease in Chinese Cabbage from Yuanyang County, Henan Province.
1 Results and Analysis
1.1 Morphological and cytological observation
The diseased Chinese cabbage plant was found with leaves wilting and root underground swelling like a fist in 8.5 cm diameter (Figure 1).
.png) Figure 1 The morphological symptoms of infected Chinese cabbage with clubroot |
After optical microscope observation, a large number of spores were found being arranged in the root cells like fish eggs and their diameters range from 2.17 µm to 5.18 µm (Figure 2), which was close to 1.9µm -4.3µm in previous report (Suo et al., 2015). The spore concentration was adjusted at about 7×107/mL.
.png) Figure 2 Cytological observation of pathogen from YY clubroot isolate in Chinese cabbage Note: Left: The observation result by using optical microscope(×400) showed that a large number of spores arranged in the root cells like fishs eggs; Right: Diameters of spores range from 2.17 µm~5.18 µm |
1.2 Pathogenicity identification
Investigation results showed that YX55 seedlings inoculated with 1mL spore suspension were obviously smaller and shorter than those without inoculation. And all roots of infected YX55 seedlings produced galls referred to as “clubs” with the shape of spindle or finger, while roots of uninfected YX55 seedlings were healthy and normal (Figure 3). The result indicated that spores isolated from the clubbed roots still had active pathogenicity.
.png) Figure 3 Pathogenicity identification result Note: Left: clubroots of inoculated seedlings after 45d; Right: healthy roots of seedlings without inoculation after 45d |
1.3 D85819 sequence analysis
YY isolate DNA sample from clubroot-infected roots gives rise to a single band with the target size of approximate 800 bp using the P. brassicae D85819-specific primers PBF6/PBR6, while no bands both in DNA from healthy B.rapa root and the control water (Figure 4). After sequenced and submitted to GenBank, the blast results showed that the target sequence shared 99% identity with D85819 sequence, a genomic survey sequence of P. brassicae from GenBank.
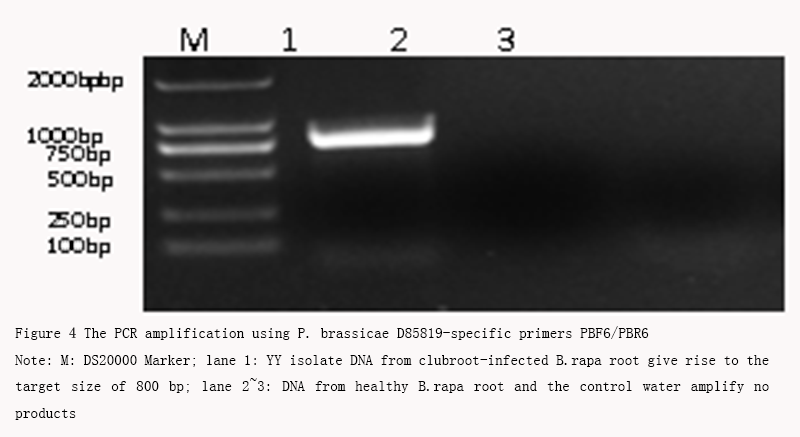 Figure 4 The PCR amplification using P. brassicae D85819-specific primers PBF6/PBR6 Note: M: DS20000 Marker; lane 1: YY isolate DNA from clubroot-infected B.rapa root give rise to the target size of 800 bp; lane 2~3: DNA from healthy B.rapa root and the control water amplify no products |
1.4 Pro1 gene sequence analysis
To further study the possibility of application the putative pathogenic-related Pro1 gene to P. brassicae identification, PCR amplification was conducted using the P. brassicae Pro1-specific primers yPro1E1F1/ yPro1E9R2. The result showed that YY isolate DNA sample from clubroot-infected root yields a single band with the target size of approximate 1,700 bp while no bands both in DNA from healthy B.rapa root and the control water (Figure 5), which demonstrated the same result with D85819 amplification.
.png) Figure 5 The PCR amplification using P. brassicaepro1-specific primers yPro1E1F1/ yPro1E9R2 Note: M: DS20000 Marker; lane 1: YY isolate DNA from clubroot-infected B.rapa root give rise to the target size of about 1,700 bp; lane 2 and 3: DNA from healthy B.rapa root and the control water amplify no products |
After sequenced and submitted to GenBank, the blast results showed that the target sequence shared 93% identity with Pro1 gene from Genbank.
In summary, by morphological, cytological observation and pathogenicity identification as well as molecular marker, the pathogen was finally identified as P. brassicae Woron.
2 Discussion
With the cultivation areas spreading wider and wider, more and more areas were found clubroot disease caused by P. brassicae unexpectedly (Chen and Wang, 2015). The pathogen in Yuanyang County was finally identified as P. brassicae Woron. No reports about P. brassicae were reported there before. Given that the local cultivation field is mostly saline-alkali land with higher pH value, and the seedlings were cultivated by substrate seedling nursing, there is a strong possibility that the pathogen were brought to Yuanyang County with the substrate bought.
For further research, DNA of P. brassicae in the same substrate should be extracted and amplified to be compared with that extracted from clubbed roots of the diseased seedling. Moreover, to find the infecting mechanism under such an unideal condition, further research about P. brassicae itself should be done. Laila et al. collected 11 Korean field populations of P. brassicae from 9 different locations and studied rDNA sequences of them, then sequence variations were observed in both the SSU (small subunit) and LSU (large subunit) of rDNA. It was considered that variation in the SSU of P. brassicae rDNA is associated with evolutionary changes that result in distinct field isolates. And mutations in rDNA sequence might influence the functionality of this protist (Laila et al., 2017). In the subsequent research, different geographic isolates in China should be collected and sequenced to identify the Pro1 gene sequence diversity among them. Since P. brassicae from different sources or different isolates show different infection capacity, it was inferred that one or several genes may play a key role in infection progress. Several important genes in P. brassicae such as GH3, PbBSMT as well as Pro1 mentioned above should be studied on different isolates of P. brassicae to find if they were related to pathogenicity diversity of P. brassicae.
As PCR-based assays have been the most effective and accurate methods for detection of P. brassicae. Much more P. brassicae-specific markers should be developed. D85819 sequence and Pro1 gene sequence in the experiment could be recommended as an effective marker to detect the pathogen. However, resulting from the development of environment and the evolutionary changes along with the spread of P. brassicae, sequences might represent diversity, which indicated that new markers might be developed to distinguish a specific isolate from the others.
Last but not the least, much more attention should still be paid to breeding. Even many strategies have been proposed for clubroot disease control, but the use of resistant cultivars is still believed to be most effective (Peng et al., 2014; Zhang et al., 2015). The experiment result here may lay a foundation to further research about P. brassicae.
3 Materials and Methods
3.1 Plant materials
The diseased Chinese cabbage seedling named “Y914” was collected and stored at -20°C. It was named “YY” for the subsequent experiment.
3.2 Designing specific primers
Primer pair PBF6/PBR6 was designed based on genomic DNA sequence (D85819) of P. brassicae, while the other primer pair yPro1E1F1/yPro1E9R2 was designed based on Pro1 gene of P. brassicae. The primers were both designed by using Primer Premier 5.0 and synthesized by Beijing Genomics Institution (BGI), with information listed in Table 1. PCR buffer and Taqase used were from Vazyme Biotech Company and dNTPs from Generay Biotech Company.
.png) Table 1 Primer sequence of D85819 and Pro1gene used in this experiment |
3.3 Preparation of resting spores
For plant inoculation, the pathogen resting spores were prepared as follows: (1) 3.5 g of fresh and clean root sample was cut into pieces and divided into 20 2 mL-centrifuge tubes, with about 0.15 g in each tube. After that, a steel bead and 500 µL sterilized ddH2O was added in each tube; (2) After crushed, these slurry was filtrated and suspension was collected into an 50 mL centrifuge tube. Then some sterilized ddH2O was added to wash the tube and eventually set the final volume at 16mL; (3) Suspension was divided into two 15 mL equally and centrifuged at 4,000 r/min for 15 min, then the supernatants were removed; (4) The pellet of each tube was re-suspend in 5 mL 50% (w/v) sucrose by gentle pipetting. After centrifugation at 4,000 r/min for 5 min, the supernatant was transferred into a new tube containing 3mL sterilized ddH2O; (5) After centrifugation at 4,000 r/min for 5 min, supernatant was removed gently; (6) Pellet was re-suspended in 6 mL sterilized ddH2O and centrifuged at 4,000 r/min for 2 min (repeat this operation for one more time); (7) The resultant pellet was re-suspended in 6mL sterilized ddH2O, and after combining and adjusting the spore concentration, the resting spore suspension was ready for the following experiment.
3.4 Plant inoculation
Indoor inoculation experiment was conducted to confirm if the pathogen spores still had active infectivity. After germination of 30 YX55 seedlings, a kind of clubroot-susceptible Chinese cabbage, 20 of them were selected and transfered into 20 plastic pots containing enough sterilized substrate for growing. 10 seedlings were inoculated with 1mL 7×107/mL resting spore suspension prepared, while the other 10 seedlings were not inoculated. All 20 pots of seedlings were placed in a greenhouse under the same conditions. Keep proper temperature and humidity. Investigation was conducted in 45 days after inoculation.
3.5 Genomic DNA extraction
Genomic DNA was extracted from clubbed roots as well as healthy root as control mainly using 2% CTAB protocol described by Liang Bao-ping et al (Liang et al., 2012). The improved protocol was as follows: (1) cut off a small block of clubroot and chopped it up into pieces, then put them into a 2 mL Eppendorf tube containing a steel bead, then added 1 mL 2% CTAB into the tube; (2) after crushing by instrument, put the tube into a water bath at 65°C for 1 h; (3) cool down them to room temperature, then add 500 µL chloroform and mix them completely, centrifuge at 12,000 r/min for 10 min; (4) transfer the supernatant into another 1.5 mL tube and add 300 µL isopropanol into it. Then centrifuge at 12,000 r/min for 20sec; (5) remove the supernatant and wash the pellet with 75% ethanol, then centrifuge at 12,000 r/min for 2 min; (6) remove the supernatant gently and dry by airing in the room. When dried, add 100 µL sterilized ddH2O into the tube to dissolve the DNA pellet. DNA was analyzed by 1% agarose gel electrophoresis and G-box gel imaging system, and the concentration of it was measured by Multiscan GO from Thermo. With DNA concentration determined at 50 ng/µL, final DNA solution was stored at -20°C for subsequent use.
3.6 PCR amplification
PCR were conducted using mentioned primers with a final volume of 50µL system as follows: 5 µL 10×Taq buffer (Mg2+ included), 1 µL dNTPs (10 mmol/L), 2 µL forward and 2µL reserve primer (10 µmol/L), 0.25 µL Taq DNA Polymerase (5 U/µL), 10 µL 10×DNA , 29.75 µL ddH2O to make system up to 50 µL. For PCR program, the denaturing time was 45 s or 60 s, the annealing temperature was 58°C for both primers, the annealing time was 45 s or 60 s, the elongation time was 60 s or 90s, and the cycle number was 35. PCR products were analyzed by 1% agarose gel electrophoresis and G-box gel imaging system.
3.7 Sequence analyses
Target PCR fragment was retained by GO-GELS-2500 kit (Gene On Bio Tech, China), connected with pMD18-T vector (Takara, Dalian) and transformed to competent cell E. Coli DH5α (Takara, Dalian). Then 3 monoclones were sent to Beijing Genomics Institution Limited Company for sequence analysis.
Author’s contributions
Zhang Xiaowei and Tian Baoming conceived and designed the experiments. Li Yang and Yuan Yuxiang performed the experiments. Wei Xiaochun designed primers. Zhao Yanyan, Yao Qiuju, Wang Zhiyong, Yang Shuangjuan and Jiang Wusheng collected samples and participated in experiments and sequence alignment. Li Yang drafted the manuscript. Yuan Yuxiang, Wei Xiaochun and Zhang Xiaowei revised the manuscript. All authors read and approved the final version of manuscript.
Acknowledgements
This work was supported by the project of National Key Research and Development Program of China (2016YFD0100204-18), Henan provincial science and technology open cooperation project (162106000011), the Excellent Young Scientist Foundation of Henan Academy of Agricultural Sciences (2016YQ11), the Specialized Scientific Research Fund of Henan Academy of Agricultural Sciences (20157805), the Natural Science Foundation of Henan Province (162300410162), and Scientific observing and experimental station of vegetable (Henan Huanghuai region), ministry of Agriculture (10205020).
Chen X., and Wang C., 2015, Pathogen identification of cabbage clubroot in Harbin Area, Plants:basel, Switzerland, 2(4): 726-749
Dixon G.R., 2009, The Occurrence and Economic Impact of Plasmodiophora brassicae and Clubroot Disease, Journal of Plant Growth Regulation, 28(3): 194-202
https://doi.org/10.1007/s00344-009-9090-y
Faggian R., and Strelkov S.E., 2009, Detection and Measurement of Plasmodiophora brassicae, Journal of Plant Growth Regulation, 28(3): 282-288
https://doi.org/10.1007/s00344-009-9092-9
Feng J., Hwang R., Hwang S.F., Strelkov S.E., Gossen B.D., Zhou Q.X., and Peng G., 2010, Molecular characterization of a serine protease Pro1 from Plasmodiophora brassicae that stimulates resting spore germination, Mol Plant Pathol, 11(4): 503-512
https://doi.org/10.1111/j.1364-3703.2010.00623.x
Hwang S.F., Strelkov S.E., Feng J., Gossen B.D., and Howard R.J., 2012, Plasmodiophora brassicae: a review of an emerging pathogen of the Canadian canola (Brassica napus) crop, Mol Plant Pathol, 13(2): 105-113
https://doi.org/10.1111/j.1364-3703.2011.00729.x
Ito S., Maehara T., Maruno E., Tanaka S., KameyaIwaki M., and Kishi F., 2008, Development of a PCR-based assay for the detection of Plasmodiophora brassicae in soil, Journal of Phytopathology, 147(2): 83-88
https://doi.org/10.1111/j.1439-0434.1999.tb03812.x
Ito S., Maehara T., Tanaka S., Kameyaiwaki M., Yano S., and Kishi F., 1997, Cloning of a single-cope DNA sequence unique to Plasmodiophora brassicae, Physiological & Molecular Plant Pathology, 50(5): 289-300
https://doi.org/10.1006/pmpp.1997.0087
Karling J.S., 1968, The Plasmodiophorales, 2nd edn, Hatner: New York
Kim W.G., Moon M.H., Kim J.H., Choi H.W., and Hong S.K., 2009, Occurrence of Clubroot on Pak-Choi Caused by Plasmodiophora brassicae, Mycobiology, 37(1): 69-71
https://doi.org/10.4489/MYCO.2009.37.1.069
PMid:23983511 PMCid:PMC3749460
Laila R., Robin A.H., Yang K., Choi G.J., Park J.I., and Nou I.S., 2017, Detection of Ribosomal DNA Sequence Polymorphisms in the Protist Plasmodiophora brassicae for the Identification of Geographical Isolates, Int J Mol Sci, 18(84): 1-16
https://doi.org/10.3390/ijms18010084
Li Y., 2010, Diagnose and control technology of Plasmodiophora brassicae, Thesis for M.S., Northeast Agricultural University, Supervisor: Xiang W.S., 11
Li Z., Niu Y.Z., Wu Y.C., and Guo S.X., 2013, Research Advances in Clubroot Disease, Southwest China Journal of Agricultural Sciences, 26(4): 1733-1737
Liang B.P., Yuan Y.X., Piao F.Z., Zhang X.W., Jiang W.S., Liang S., Yao Q.J., Zhang Q., and Zhao Y.Y., 2012, The optimization of technique of native polyacrylamide gel electrophoresis (Native-PAGE), Journal of Henan Agricultural Sciences, 41(5): 129-132
Niwa R., Kawahara A., Murakami H., Tanaka S., and Ezawa T., 2011, Complete structure of nuclear rDNA of the obligate plant parasite Plasmodiophora brassicae: intraspecific polymorphisms in the exon and group I intron of the large subunit rDNA, Protist, 162(3): 423-434
https://doi.org/10.1016/j.protis.2011.02.005
PMid:21497131
Peng G., Lahlali R., Hwang S.F., Pageau D., Hynes R.K., Mcdonald M.R., Gossen B.D., and Strelkov S.E., 2014, Crop rotation, cultivar resistance, and fungicides/biofungicides for managing clubroot (Plasmodiophora brassicae) on canola, Canadian Journal of Plant Pathology, 36(sup1): 99-112
Suo H., Chen L., Xu H., Song B., Fan X., Yuan X., and He C., 2015, Research progress in clubroot of crucifers, Journal of Anhui Agricultural Sciences, (14): 115-117
Wallenhammar A.C., 1996, Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels, Plant Pathology, 45(4): 710-719
https://doi.org/10.1046/j.1365-3059.1996.d01-173.x
Wallenhammar A.C., and Arwidsson O., 2001, Detection of Plasmodiophora Brassicae By PCR in Naturally Infested Soils, European Journal of Plant Pathology, 107(3): 313-321
https://doi.org/10.1023/A:1011224503200
Wang J., Huang Y., Xiao-Ling H.U., Niu Y.Z., Xiao-Lan L.I., and Liang Y., 2008, Study on symptom, yield loss of clubroot and modality of Plasmodiophora brassicae in rape, Chinese Journal of Oil Crop Sciences, 30(1): 112-115
Yang P.W., Yang Q.Z., Wang Q., Li J.R., and Zeng L., 2002, PCR detection of Plasmodiophora brassicae causing cruciferae clubroot, Journal of Yunnan Agricultural University, 17(2): 137-139
Yang S.C., 2013, Study on resistance of different cabbage and Chinese variaties and detection of Plasmodiophora brassicae in soils, Thesis for M.S., Southwest University, Supervisor: Li C.Q.pp.28-37
Zhang H., Feng J., Manolii V.P., Strelkov S.E., and Hwang S.F., 2015, Characterization of a Gene Identified in Pathotype 5 of the Clubroot Pathogen Plasmodiophora brassicae, Phytopathology, 105(6): 764-770
https://doi.org/10.1094/PHYTO-10-14-0270-R
PMid:25689519



. PDF(519KB)
. FPDF(win)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Yang Li
. Yuxiang Yuan
. Yanyan Zhao
. Xiaochun Wei
. Qiuju Yao
. Wusheng Jiang
. Zhiyong Wang
. Shuangjuan Yang
. Xiaowei Zhang
. Baoming Tian
Related articles
. Yuanyang County
. Clubroot disease
. Chinese cabbage
. Pathogen indentification
Tools
. Email to a friend
. Post a comment